Hoy les traigo el descubrimiento más reciente sobre la historia evolutiva de los manatíes. Y es que en esta nueva publicación científica junto con los colegas Catalina Suarez, Javier N. Gelfo y Jorge W. Moreno-Bernal, describimos los restos más antiguos de manatíes de las Américas. En la entrada previa de esta serie ya les había adelantado lo que sabíamos hasta el momento (2015) sobre la historia evolutiva de los manatíes, incluyendo el origen europeo del grupo. En este nuevo trabajo describimos una porción de un maxilar izquierdo con las dos muelas más posteriores (denominadas como M2 y M3), proveniente de la Formación Barzalosa, la cual se depositó entre 16.5-17.7 millones de años, lo cual nos ayuda a entender mejor los orígenes de las especies actuales de manatí (Suarez et al., 2021). Al estudiar y examinar el fósil de la Formación Barzalosa llegamos a la conclusión que guarda mucha similitud con Potamosiren magdalenensis, una especie de manatí extinto que vivía en lo que hoy día es Colombia entre 11.6-13.8 millones de años atrás (Reinhart, 1951; Kellogg, 1966; Domning, 1997). Por su parecido, designamos a este nuevo fósil como Potamosiren cf. P. magdalenensis, y pasa a ser el registro más antiguo de esta especie. Algo que distingue a Potamosiren de otros sirenios, especialmente de otros manatíes, son dos características particulares de sus dientes. La primera es que todavía retienen tres molares por cada cuadrante y la segunda, es que el esmalte de las muelas es inusualmente grueso.
A-C. Maxilar izquierdo de
Potamosiren cf.
P. magdalenensis de la Formación Barzalosa.
D. Maxilar izquierdo de
Potamosiren magdalenensis del Grupo Honda. (Modificado de Suarez et al., 2021).
El
Potamosiren de la Fm. Barzalosa, junto a una especie poco conocida de manatí del Mioceno temprano de Perú (Antoine et al., 2016), representan los restos más antiguos de manatíes en las Américas. Ambos, coinciden geográficamente y temporalmente con el inicio de la formación del Mega-Humedal de Pebas, el cual abarcó una enorme zona entre 23-10 millones de años, e incluía partes de Perú, Colombia y Brazil (Hoorn et al., 2010). Este mega-humedal estaba conectado con el mar Caribe hacia el norte, lo cual funcionó como vía de entrada para distintos organismos marinos. Los depósitos sedimentarios que se formaron durante la duración de el Mega-Humedal de Pebas están principalmente compuestos de estratos que representan ambientes terrestres y de agua dulce, con algunas incursiones marinas. Mientras que el manatí de Pebas viene de sedimentos que representan ambientes acuáticos mixtos, los del
Potamosiren de Barzalosa son de agua dulce, ambos entonces representan los primeros registros de sirenios del grupo terminal (crown group) fuera de ecosistemas marinos. (En este contexto,
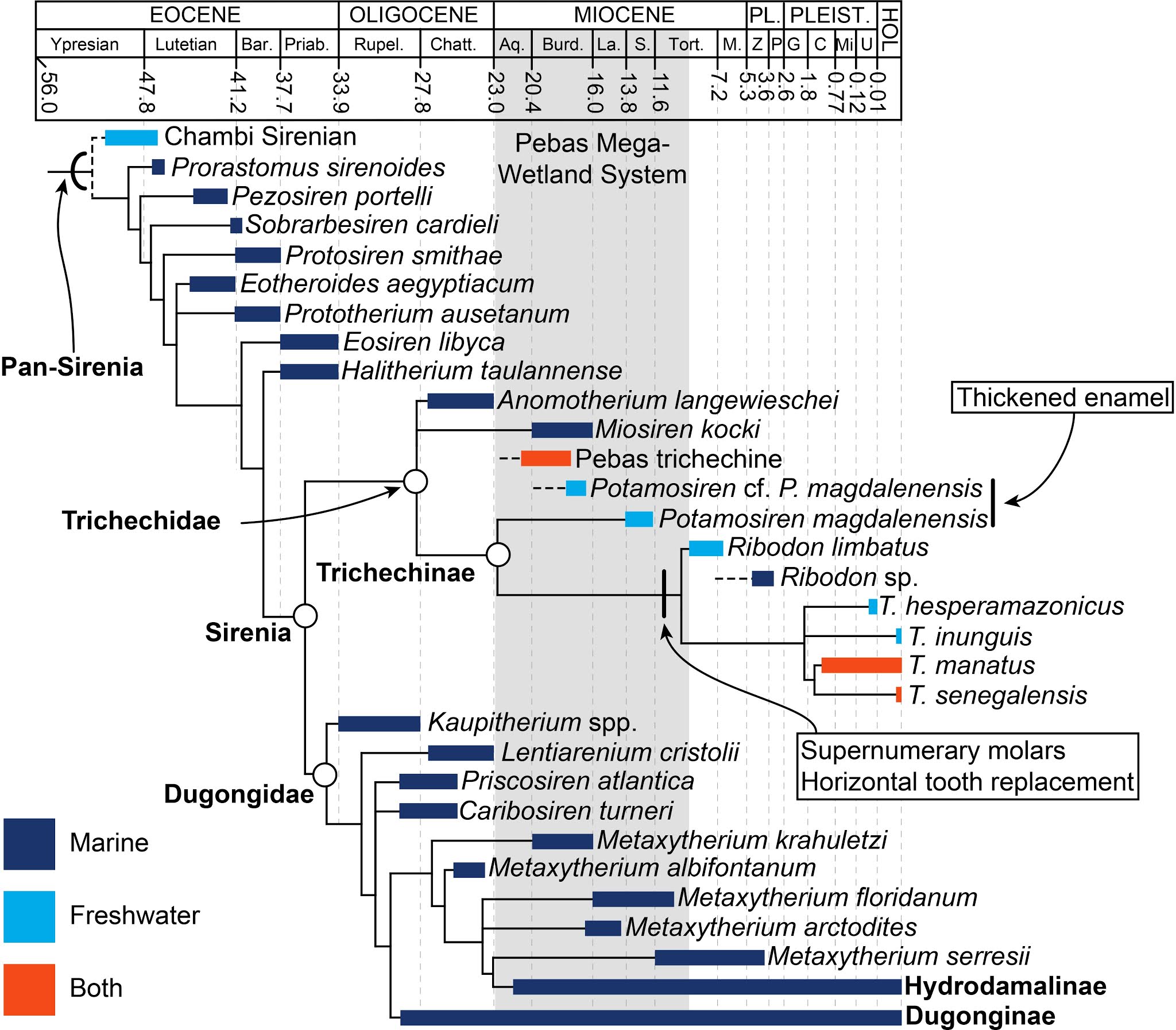
Sirenia es el crown group, que es el grupo más exclusivo compuesto de las especies actuales y sus parientes más cercanos, y
Pan-Sirenia es el grupo total que incluye las especies extintas basales y las actuales; ver imagen abajo).
En este árbol filogenético se puede apreciar las relaciones entre los distintos grupos de sirenios, con énfasis en los manatíes (Trichechidae). En gris está representado la duración del Mega-Humedal de Pebas y los distintos colores representan los ambientes donde se encuentran las distintas especies. (Modificado de Suarez et al., 2021).
Como se puede observar en el árbol (ver arriba) entre los sirenios, solamente los manatíes (Trichechinae) incursionan en ambientes de agua dulce. Esto contrasta con otros grupos de organismos marinos. Por ejemplo, los odontocetos han incursionado en ambientes de agua dulce en múltiples ocasiones y en diversas partes del mundo (Fordyce, 1983; Cassens et al., 2000; Hulbert & Whitmore, 2006; Geisler et al., 2011; Boessenecker & Poust, 2015; Bianucci et al., 2013; Pyenson et al., 2015; Boersma & Pyenson, 2016; Benites-Palomino et al., 2020). El Mega-Humedal de Pebas no fue la excepción ya que se han encontrado fósiles de
iniidos y
platanistidos en distintas localidades (Bianucci et al., 2013; Benites-Palomino et al., 2020).
América del Sur y Central alrededor de 12 millones de años atrás, mostrando las localidades donde se han encontrado iniidos y platanistidos en ambientes marinos (azul) y de agua dulce (amarillo). (Modificado de Benites-Palomino et al., 2020).
En adición a los mamíferos marinos, el Mega-Humedal de Pebas fue un lugar excepcional para que distintos grupos de organismos de origen marino incursionaran y se adaptaran a ambientes de agua dulce y pantanosos, como por ejemplo las mantarrayas de agua dulce (Fontenelle et al., 2021) y algunos otros grupos de peces (Bloom & Lovejoy, 2017). Incluso algunos grupos se diversificaron para ocupar distintos espacios ecológicos y proliferaron durante la duración de este mega-humedal (Salas-Gismondi et al., 2015). No es la primera vez que se propone que la formación del Mega-Humedal de Pebas haya sido clave para la incursión de manatíes en cuerpos de agua dulce. Esta hipótesis se propuso por primera vez en los 1980's (Domning, 1982) y más recientemente en un estudio genético de las especies actuales (Silva de Zouza et al., 2021). El
Potamosiren de Barzalosa es evidencia fósil que le da aún más veracidad a esta hipótesis (Suarez et al., 2021).
Las muelas de PotamosirenDiferentes grupos de mamíferos herbívoros han evolucionado distintas adaptaciones como resultado de tener una dieta que incluye vegetación abrasiva o fibrosa. Por ejemplo, los caballos tienen dientes hipsodontes (dientes con coronas muy altas) que los hacen más resistentes o duraderos al desgaste. Otra posible alternativa para contrarrestar el desgaste es que el esmalte del diente sea más grueso de lo usual, haciéndolo más resistente. Mientras que una tercera estrategia es tener dientes que pueden ser reemplazados a medida que se van gastando.Como mencione previamente, el esmalte inusualmente grueso de las muelas de
Potamosiren es único entre los sirenios. Sin embargo, no es la primera vez que se observa este fenómeno en mamíferos marinos herbívoros. Y es que el esmalte engrosado nos recuerda a las muelas de
Desmostylus, un mamífero marino herbívoro que habitaba a lo largo de las costas del Pacífico Norte hasta hace unos 10-12 millones de años.
Desmostylus pertenece a un grupo completamente extinto llamado Desmostylia, y sus cráneos y esqueletos son tan particulares que todavía queda mucho por descifrar sobre su paleoecología e incluso sobre su postura en tierra y su relación con otras especies. Sin embargo, gracias a estudios isotópicos usando los dientes de
Desmostylus conocemos un poco sobre su dieta (Clementz et al., 2003). Los resultados de ese estudio sugieren que los
Desmostylus consumían distintos tipos de vegetación acuática, algunas, como los pastos marinos, cuales inevitablemente debía incluir interacción con el substrato e ingestión accidental de sedimento; en fin, una dieta relativamente variada y muy parecida a la del manatí de Florida (
Trichechus manatus latirostris), los cuales tienen una dentadura muy particular como veremos más adelante. Como nota al calce, hay que mencionar que la dieta de los manatíes actuales es una excepción, ya que los sirenios han sido casi exclusivamente consumidores de pastos marinos durante la mayor parte de su historia evolutiva (Clementz, et al., 2003; MacFadden et al., 2004; Clementz & Sewall, 2011), y su morfología dental ha sido generalmente muy conservadora. Así que cuando observamos cualquier cambio significativo en la dentadura de una especie de sirenio inmediatamente despierta la curiosidad por conocer mejor sus hábitos alimenticios.
Parte de la mandíbula de un
Desmostylus, incluyendo el cuarto premolar y las primeras dos muelas. Las muelas están compuestas de cúspides modificadas en columnas con esmalte muy grueso.
Los distintos ecosistemas que formaban parte del Mega-Humedal de Pebas permitieron que los manatíes tuviesen acceso a una cantidad mayor de vegetación, en adición a un incremento en la interacción con vegetación acuática cerca del substrato e ingestión accidental de sedimento. Esto claramente debió resultar en un cambio significativo en la dieta de estos sirenios (Domning, 1982; Beatty et al., 2012), que como antes mencionado, eran principalmente consumidores de pastos marinos. Tomando esto en consideración, en nuestro trabajo proponemos que el esmalte engrosado de las muelas de
Potamosiren es una adaptación a una dieta más variada, incluyendo mayor cantidad de vegetación abrasiva y fibrosa. Y como verán, las especies de manatí que vinieron después de
Potamosiren se adaptaron a esta dieta de una forma aún más particular.
Molares superiores (izquierda) y mandíbula (derecha) de
Potamosiren magdalenensis. Aquí se puede apreciar la presencia de solamente tres molares por cuadrante.
Posterior a la extinción de
Potamosiren aparece otra especie de manatí conocido como
Ribodon limbatus el cual también se encuentra en depósitos de agua dulce del Mioceno tardío al Plioceno.
Ribodon se diferencia de
Potamosiren al tener más de tres molares por cuadrante (usualmente entre 4-5), los cuales son muy similares entre sí y son relativamente pequeños, tampoco tienen esmalte engrosado.
Ribodon también se caracteriza porque sus muelas eran reemplazadas gradualmente. En otras palabras, a lo largo de la vida de un individuo las muelas se movían lentamente hacia al frente mientras que en la parte posterior iban saliendo muelas nuevas, una y otra vez! Este tipo de reemplazo horizontal de los dientes es una adaptación que se observa por primera vez en
Ribodon y que todavía observamos en las tres especies de manatí actuales.
Detalles de la dentición de
Ribodon (arriba) y
Trichechus (abajo). Noten la presencia de multiples muelas, de similar morfología y tamaño, al igual que los dientes nuevos hacia la parte posterior de la fila dental.
Los manatíes actuales pertenecen al género
Trichechus, e incluyen tres especies -
T. manatus (manatí antillano),
T. inunguis (manatí amazónico) y
T. senegalensis (manatí africano). En adición a estas tres especies, se conoce una subespecie extinta del manatí antillano del Pleistoceno de Norteamérica (
T. manatus bakerorum) y una especie del Pleistoceno amazónico (
T. hesperamazonicus) (Domning, 2005; Perini et al., 2020). Todas las especies de
Trichechus (actuales y extintas) reemplazan sus muelas horizontalmente, y estas son aún más pequeñas que las de
Ribodon, teniendo en algunos casos hasta siete muelas por cuadrante! Esto resulta en un sistema relativamente eficiente donde se reemplazan los dientes continuamente para contrarrestar una dieta variada que puede incluir vegetación fibrosa e ingestión accidental de sedimento. Sin embargo, no es perfecto, y la composición y tipo de sedimento también juega un papel importante. En lugares como el estado de Florida donde la arena tienen una mayor composición de cuarzo, el desgaste en los dientes de los manatíes es mayor que el observado en los que viven en el Caribe, donde las arenas están principalmente compuestas de carbonato de calcio (Domning & Hayek, 1984).
Aún con esta nueva información, queda mucho por conocer sobre la historia evolutiva de los manatíes. Por ejemplo, todavía no conocemos bien la identidad del manatí de Pebas y tampoco conocemos bien la morfología craneal de
Potamosiren y
Ribodon. Similarmente, faltan estudios isotópicos que nos puedan dar una mejor idea de la dieta de estas especies de manatíes ancestrales. Esperemos que tengamos contestaciones a estas incógnitas en un futuro no tan lejano.
Literatura
Antoine, P.-O., M. A. Abello, S. Adnet, A. J. Altamirano Sierra, P. Baby, G. Billet, M. Boivin, Y. Calderón, A. Candela, J. Chabain, F. Corfu, D. A. Croft, M. Ganerod, C. Jaramillo, S. Klaus, L. Marivaux, R. E. Bavarrete, M. J. Orliac, F. Parra, M. E. Pérez, F. Pujos, J. C. Rage, A. Ravel, C. Robinet, M. Roddaz, J. V. Tejada-Lara, J. Velez-Juarbe, F. P. Wesselingh, and R. Salas-Gismondi. 2016. A 60-million-year Cenozoic history of western Amazonian ecosystems in Contamana, eastern Peru. Gondwana Research 31:30–59.
Beatty, B. L., T. Vitkovski, O. Lamnert, and T. E. Macrini. 2012. Osteological associations with unique tooth development in manatees (Trichechidae, Sirenia): a detailed look at modern Trichechus and a review of the fossil record. Anatomical Record 295:1504–1512.
Benites-Palomino, A., G. Aguirre-Fernandez, J. W. Moreno-Bernal, A. Vanegas, and C. Jaramillo. 2020. Miocene freshwater dolphins from La Venta, Huila, Colombia suggest independent invasions of riverine environments in tropical South America. Journal of Vertebrate Paleontology 40:e1812078.
Bianucci, G., O. Lambert, R. Salas-Gismondi, J. Tejada, F. Pujos, M. Urbina, and P.-O. Antoine. 2013. A Miocene relative of the Ganges river dolphin (Odontoceti, Platanistidae) from the Amazonian basin. Journal of Vertebrate Paleontology 33:741–745.
Bloom, D. D., and N. R. Lovejoy. 2017. On the origins of marine-derived freshwater fishes in South America. Journal of Biogeography 44:1927–1938.
Boersma, A. T., and N. D. Pyenson. 2016. Arktocara yakataga, a new fossil odontocete (Mammalia, Cetacea) from the Oligocene of Alaska and the antiquity of Platanistoidea. PeerJ 4:e2321.
Boessenecker, R. W., and A. W. Poust. 2015. Freshwater occurrence of the extinct dolphin Parapontoporia (Cetacea: Lipotidae) from the upper Pliocene nonmarine Tulare Formation of California. Palaeontology 58:489–496.
Cassens, I., S. Vicario, V. G. Waddell, H. Balchowsky, D. Van Belle, W. Ding, C. Fan, R. S. Lal Mohan, P. C. Simoes-Lopes, R. Bastida, A. Meyer, M. J. Stanhope, and M. C. Milinkovitch. 2000. Independent adaptation to riverine habitats that allowed survival of ancient cetacean lineages. Proceeding of the National Academy of Sciences U.S.A. 97:11343–11347.
Clementz, M. T., and J. O. Sewall. 2011. Latitudinal gradients in greenhouse seawater d18O: evidence from Eocene sirenian tooth enamel. Science 332:455–458.
Clementz, M. T., K. A. Hoppe, and P. L. Koch. 2003. A paleoecological paradox: the habitat and dietary preferences of the extinct tethythere Desmostylus inferred from stable isotope analysis. Paleobiology 29:506–519.
Domning, D. P. 1982. Evolution of manatees: a speculative history. Journal of Paleontology 56:599–619.
Domning, D. P. 1997. Sirenia. In: R. F. Kay, R. H. Madden, R. L. Cifelli, and J. J. Flynn (Eds.), Vertebrate Paleontology in the Neotropics: the Miocene Fauna of La Venta, Colombia. Smithsonian Institution Press, Washington and London, pp. 383–391.
Domning, D. P., and L.-A. C. Hayek. 1984. Horizontal tooth replacement in the Amazonian manatee (Trichechus inunguis). Mammalia 48:105–127.
Fontenelle, J. P., F. P. Luna Marques, M. A. Kolmann, and N. R. Lovejoy. 2021. Biogeography of the neotropical freshwater stingrays (Myliiobatiformes: Potamotrygoninae) reveals effects of continent-scale paleogeographic change and drainage evolution. Journal of Biogeography 00:1–14.
Fordyce, R. E. 1983. Rhabdosteid dolphins (Mammalia: Cetacea) from the middle Miocene, lake Frome area, South Australia. Alcheringa 7:27–40.
Geisler, J. H., M. R. McGowen, G. Yang, and J. Gatesy. 2011. A supermatrix analysis of genomic, morphological, and paleontological data from crown Cetacea. BMC Evolutionary Biology 11:1–33.
Hoorn, C., F. P. Wesselingh, H. T. Steege, M. A. Bermudez, A. Mora, J. Sevink, I. Sanmartín, A. Sanchez-Meseguer, C. L. Anderson, J. P. Figuerido, C. Jaramillo, D. Riff, F. R. Negri, H. Hooghiemstra, J. Lundberg, T. Stadler, T. Särkinen, and A. Antonelli. 2010. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 330:927–931.
Hulbert, R. C., and F. C. Whitmore. 2006. Late Miocene mammals from the Mauvilla local fauna, Alabama. Bulletin of the Florida Museum of Natural History. 46:1–28.
Kellogg, R. 1966. Fossil marine mammals from the Miocene Calvert Formation of Maryland and vVirginia, parts 3 and 4. Bulletin of the U. S. National Museum 247:65–101.
MacFadden, B. J., P. Higgins, M. T. Clementz, and D. S. Jones. 2004. Diets, habitat preferences, and niche differentiation of Cenozoic sirenians from Florida: evidence from stable isotopes. Paleobiology 30:297–324.
Pyenson, N. D, J. Velez-Juarbe, C. S. Gutstein, H. Little, D. Vigil, and A. O'Dea. 2015. Isthminia panamensis, a new fossil inioid (Mammalia, Cetacea) from the Chagres Formation of Panama and the evolution of 'river dolphins' in the Americas. PeerJ 2015: e1227.
Reinhart, R. H. 1951. A new genus of sea cow from the Miocene of Colombia. Bulletin of the Department of Geological Sciences 28:203–214.
Salas-Gismondi, R., J. J. Flynn, P. Baby, J. V. Tejada-Lara, F. P. Wesselingh, and P.-O. Antoine. 2015. A Miocene hyperdiverse crocodylian community reveals peculiar trophic dynamics in proto-Amazonian mega-wetlands. Proceedings of the Royal Society B 282:20142490.
Silva de Souza, E. M., L. Freitas, E. K. da Silva Ramos, G. Selleghin-Veiga, M. C. Rachid-Ribeiro, F. A. Silva, M. Marmontel, F. Rodrigues dos Santos, A. Laudisoit, E. Verheyen, D. P. Domning, and M. Freitas Nery. 2021. The evolutionary history of manatees told by their mitogenomes. Scientific Reports 11:3564.
Suarez, C., J. N. Gelfo, J. W. Moreno-Bernal and J. Velez-Juarbe. 2021. An early Miocene manatee from Colombia and the initial Sirenian invasion of freshwater ecosystems. Journal of South American Earth Sciences 109:103277







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}